

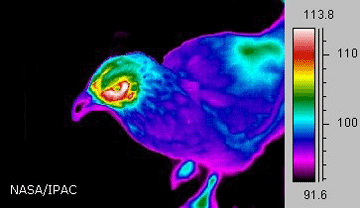
Рис. 1.1.1.1. Зависимость величины теплоотдачи
от плотности поверхностного покрова
у разных видов животных
(температура на шкале выражена в градусах Фаренгейта)
www.spitzer.caltech.edu
1.1.1. Терморегуляция и теплообмен организма животных с окружающей средой
Способность организма поддерживать постоянство температуры на определенном уровне при изменяющихся условиях внешней среды называется терморегуляцией. В основе данного явления лежит относительное равновесие между процессами, обеспечивающими образование тепла (химическая терморегуляция) и его отдачу во внешнюю среду (физическая терморегуляция).
Химическая терморегуляция осуществляется путем изменения уровня обмена веществ и усиления или ослабления процессов теплообразования под влиянием температуры воздуха.
Основное влияние на теплопродукцию оказывает температура окружающей среды в сочетании с влажностью и движением воздуха.
Помимо химической терморегуляции тепловое равновесие в организме обеспечивается отдачей излишков тепла в окружающую среду посредством физических процессов — испарения, теплопроведения, конвекции, теплоизлучения (радиации).
На величину теплоотдачи влияет не только температура окружающей среды, но и наличие шерстного или перьевого покрова (рис. 1.1.1.1).
Рис. 1.1.1.1. Зависимость величины теплоотдачи
от плотности поверхностного покрова
у разных видов животных
(температура на шкале выражена в градусах Фаренгейта)
www.spitzer.caltech.edu
Как видно из представленных данных, теплоотдача у лошади осуществляется практически через всю поверхность тела, в отличие от овцы и курицы.
Важное значение в процессах терморегуляции имеют вазомоторная и пиломоторная реакции, а также учащение дыхания (полипноэ).
Каждый вид животного в зависимости от его возраста нормально развивается и дает максимальную продуктивность только при определенном оптимуме температуры среды, так называемой зоны теплового комфорта. Границы зоны зависят от вида и возрастной группы животных (рис. 1.1.1.2).
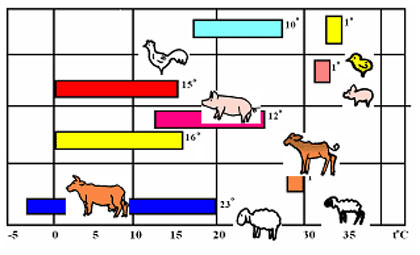
Рис. 1.1.1.2. Зоны термического комфорта у животных разного возраста и вида
(цифра, стоящая справа от каждой зоны, обозначает диапазон температуры, 0С)
В зависимости от доминирования того или иного вида терморегуляции выделяют животных с "физическим" или "химическим" типом терморегуляции (Ю.О. Раушенбах).
Под "физическим" типом терморегуляции имеется в виду такая регуляция теплообмена, при которой реакция, направленная на поддержание температурного гомеостаза, осуществляется в основном механизмами теплоотдачи. Благодаря тому, что эти механизмы обеспечивают значительное уменьшение теплоотдачи, теплопродукция не возрастает.
У животных "химического" типа, теплоотдача значительно возрастает вследствие слабого развития ее механизмов при понижении температуры среды. Благодаря этому поддержание температурного гомеостаза осуществляется за счёт повышения теплопродукции, отсюда и различия в энергетических затратах на адаптацию к изменениям температуры среды у животных "физического" и "химического" типов терморегуляции.
У видов "физического" типа терморегуляции олигопноэ и пиломоторная реакция не допускают повышение теплоотдачи и теплопродукция не возрастает.
Понижение температуры тела у животных этого типа не связана с дополнительными энергетическими затратами. Так, при понижении температуры среды от 10oС до -15oС у крупного рогатого скота, характеризуемого "физическим типом терморегуляции", интенсивность обмена или остается неизменной, или несколько снижается, тогда как у животных "химического типа терморегуляции" в этих же условиях она возрастает на 20-50%.
Следует отметить, что в аборигенных популяциях крупного рогатого скота, преобладает "физический" тип терморегуляции.
Аналогичные данные установлены и при различиях в реакциях на высокую температуру. У животных с плохо развитыми механизмами теплоотдачи, потоотделения и полипноэ адаптивная реакция на повышение температуры среды сопровождается повышением энергетических затрат, возрастает интенсивность обмена.
В противоположность этому у животных с хорошо развитыми механизмами теплоотдачи интенсивность обмена не возрастает.
Раушенбахом Ю.О. разработана формула для оценки теплоустойчивости. При составлении формулы учтена выявленная зависимость изменения температуры тела от изменения температуры среды. На основании этой зависимости рассчитан коэффициент регрессии температуры тела в зависимости от температуры среды.
Это позволяет проводить определение теплоустойчивости организма при любой температуре среды выше 30°С.
На основе этого общая формула индекса теплоустойчивости (ИТУ) преобразуется для разных видов животных следующим образом:
| Крупный рогатый скот | ИТУк=2(0,6t2 -10ΔT+26) |
|---|---|
| Овцы | ИТУо=2(0,5 t2-10ΔT+30) |
| Свиньи | ИТУс=2(0,7 t2 -10ΔT+22) |
где t2 - температура среды при температурной нагрузке;ΔT - разница в температуре тела днём при высокой температуре среды и утром в термонейтральной зоне.
Индекс холодоустойчивости может быть рассчитан по следующей формуле:
| ИХУ= | (Т2-Т1)×100 |
|---|---|
| 60-Т2К(t2 +10) |
где Т1- величина теплопродукции (ккал/ч • кг живой массы) в зоне температурного комфорта; Т2 - при низкой температуре; t2 - после двухчасовой экспозиции;
К - коэффициент регрессии изменения величины теплопродукции (в %) по понижению температуры воздуха.
Для крупного рогатого скота этот коэффициент оказался равным 0,6.
В табл. 1.1.1.1. представлены данные по различной теплоустойчивости крупного рогатого скота разных пород в условиях Средней Азии.
1.1.1.1. Породные различия в теплоустойчивости крупного рогатого скота
| Порода | ИТУ | Порода | ИТУ |
|---|---|---|---|
| Зебувидный скот | 85 | Швицкая | 70 |
| Бушуевская | 82 | Черно - пестрая | 69 |
| Красная степная | 79 | Шортгорнская | 66 |
| Англерская | 73 | Симментальская | 56 |
Зебувидный скот Средней Азии, отличающийся высокой теплоустойчивостью, имеет ИТУ, равный 85, а наименее теплоустойчивая из сравниваемых пород симментальская - 56. Породные различия по теплоустойчивости получены и по другим видам животных.
Установлено, что при высоких температурах учащение дыхание сопровождается уменьшением его глубины ( поверхностное дыхание), а повышение теплоотдачи через верхние дыхательные пути не влечёт за собой повышение теплопродукции. Такая регуляция дыхания (истинное полипноэ) становится эффективным механизмом терморегуляции при высокой температуре среды, способствуя повышению теплоустойчивости, сохранению температурного гомеостаза.
При низкой температуре, для понижения теплоотдачи рациональней положительная корреляция между этими элементами дыхания, когда при понижении частоты дыхания происходит одновременно и уменьшение его глубины (олигопноэ).
Наглядное представление о различиях в структуре реакции даёт графическое изображение относительных размеров функциональных cдвигов (рис. 1.1.1.3).
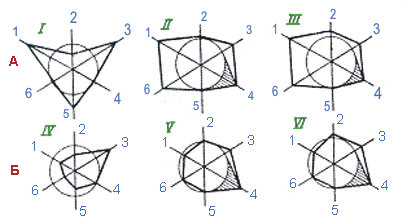
Рис. 1.1.1.3. Утрата эффективной структуры адаптивной реакции заводскими породами крупного рогатого скота:
А — на высокую температуру среды, Б — на низкую температуру среды;
Относительные размеры функциональных сдвигов (%) откладываются на векторах вне или внутри окружности, фиксирующей "норму" (100%) при нейтральных условиях среды.
Внутри окружности откладывается величина понижения, а вне окружности - величина повышения интенсивности функционирования данного механизма при воздействии фактора. Многоугольник, построенный путём соединения точек отсчёта, характеризует общую структуру реакции на высокую температуры среды у двух групп крупного рогатого скота, отличающихся по теплоустойчивости.
За 100% взяты величины, характеризующие показатели терморегуляции при 13-17°С. На векторах отложены величины по этим показателям (механизмам) при 34-37°С , в процентах "нормы" при 13-17°С.
Структура реакции на низкую температуру среды имеет определенное сходство у лошадей и крупного рогатого скота, у которых механизм физической терморегуляции направлен на уменьшение теплоотдачи.
Это достигается через сокращение объёма лёгочной вентиляции, осуществляемое посредством олигопноэ, и развития пиломоторной реакции, понижающей величину теплоотдачи с поверхности тела за счёт возрастания теплоизолирующих свойств волосяного покрова.
Оба этих механизма лучше развиты у лошади, благодаря чему при очень низких температурах - (25 - 35)°С у них не только не повышается, но зачастую, уменьшается величина теплопродукции.
У новорожденных животных, благодаря их анатомическим и физиологическим особенностям, механизмы терморегуляции функционируют несколько иначе, чем у взрослых.
Мелкие животные имеют большую поверхность тела (отдающую теплоту) по отношению к его объему (производящему теплоту). Поскольку при низких температурах с большей поверхности тела возрастает отдача тепла, молодые животные чувствительней к холоду, чем особи старших возрастов.
1.1.1. Терморегуляция и теплообмен организма животных
с окружающей средой
I — аборигенный скот Средней Азии;
II, V — черно-пестрая порода;
III— симментальская порода;
IV — аборигенный якутский скот;
VI — джерсейская порода.
1 — частота дыхания;
2 — глубина дыхания; 3 — потоотделение (при высокой температуре),
пиломоторная реакция (при низкой температуре); 4 — теплопродукция; 5 — сосудистая реакция;
6 — легочная вентиляция